Research
私たちの研究室では、「体の形がどうやって進化してきたのか」といった生命現象の本質に迫るテーマで研究を行っています。動物の形態が進化する際には、発生プログラムの変化を伴っていることから、系統的に重要な位置にいる動物種の発生プログラムを解析することで、この問題にアプローチしています。脊椎動物の進化の過程で新しく獲得され、形態を変化させてきた器官には様々なものがありますが、私たちは「手足」をモデルにして、この問題に取り組んでいます。手と足は古代魚の胸鰭と腹鰭から進化したものですが、私たちの祖先の原始脊索動物はナメクジのような形をしており、対にな�った鰭は持っていませんでした。何もないところから、鰭ができて、手足に形を変え、さらに多様な形に変わるまでに何がおきているのか?手足は体の形を進化させるメカニズムを理解する上で、とても有用なモデルです。私たちの研究室では、ニワトリ、ゼブラフィッシュ、メダカのような遺伝子改変のできるスタンダードなモデル動物に加え、必要に応じて、トラザメ、エミュー、スッポン、トビハゼ、コキコヤスガエル、、、、、など、様々な動物種をモデルに研究をすすめています。以下に私たちの研究の成果の一部を紹介します。
飛べない鳥エミューの翼の退化―胚の動きの違いが骨の形を変えるー
飛べない鳥であるエミューでは、翼が著しく退縮していますが、その形態形成のメカニズムには不明な点が多く残されていました。本研究では、エミューの翼の骨格が退縮しているだけでなく、左右非対称なパターンを示していることを見つめました。そして、その原因が翼の先端に筋肉が作られないために、胚発生中に翼をほとんど動かせず、骨の発生に必要なメカノストレスを骨になる細胞が十分に受け取れていないためであることが示されました。さらに、エミューの翼の先端に筋肉がないのは、体節由来の筋前駆細胞と側板中胚葉細胞の二重のアイデンティティを持つ筋前駆細胞が存在し、これらが融合して筋繊維になるときに、細胞死を起こすためであることを明らかにしました。本研究の成果は、胚や胎児の運動の違いが、形態進化に繋がる変化を起こす可能性があるこ�とを示唆しています。

- Eriko Tsuboi*, Satomi F. Ono*, Ingrid Rosenburg Cordeiro*, Reiko Yu, Toru Kawanishi, Makoto Koizumi, Shuji Shigenobu, Guojun Sheng, Masataka Okabe, and Mikiko Tanaka(*These authors contributed equally to this work.)(2024). Immobilization secondary to cell death of muscle precursors with a dual transcriptional signature contributes to the emu wing skeletal pattern. Nature Communications 15, 8153.
酸素で手足の形をつくるメカニズムー指間細胞死ー
カエルやイモリなどの両生類は、指や指間の成長(細胞増殖)の違いで手足の形を作っています。一方、鳥類や哺乳類などの羊膜類では、これに加えて「細胞死」によっても手足が形作られます。手足の細胞の一部が細胞死によって削り取られるようになると、手足の形は多様に進化しました。例えば、オオバンと呼ばれる水鳥は細胞死によって木の葉の形をした水かきを持つように進化し、馬やラクダは細胞死で指の数を減らしています。しかし、こうした「細胞死」による形作りのメカニズムがどのように出現したのかについては、これまで明らかにされていませんでした。私たちは、四肢動物の進化の過程で出現した指間細胞死に、驚くべき要素が必要であったことを見いだしました。大気中の「酸素」が必要だったのです。この成果は、生物の陸上進出に伴う環境の変化が、体の形成において新たなメカニズムを誕生させうることを示す重要な成果となりました。
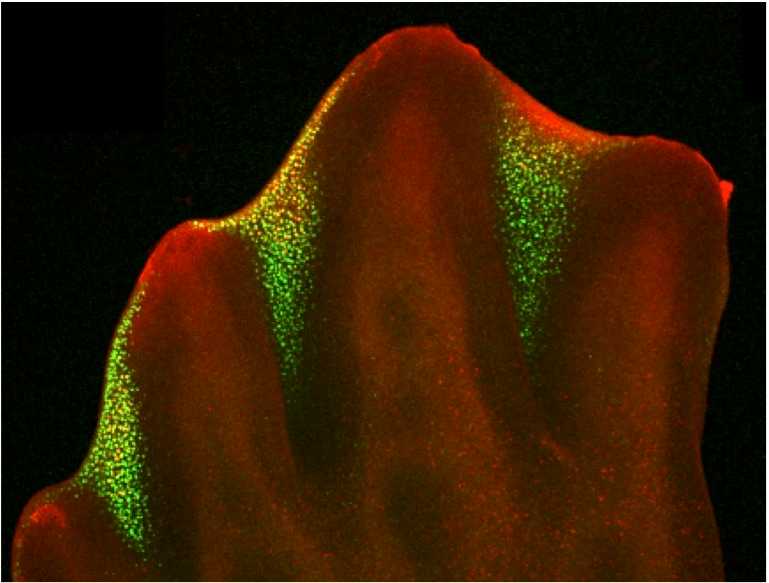
- Ingrid Rosenburg Cordeiro, Kaori Kabashima, Haruki Ochi, Keijiro Munakata, Chika Nishimori, Mara Laslo, James Hanken, and Mikiko Tanaka(2019). Environmental oxygen exposure allows for the evolution of interdigital cell death in limb patterning. Developmental Cell 50, 155-166.
手足を動かす筋肉の発生様式の進化をサメから学ぶ
四肢動物の四肢の筋肉は遊離筋とよばれる細胞によってつくられることが知られています。一方、軟骨魚類の対鰭の筋肉は、遊離筋によらない方法でつくられるとされ、四肢の筋肉の原始的な発生様式であると考えられてきました。本研究では、顎口類の原始的な状態を反映するとされる軟骨魚類において、その対鰭の筋肉の発生様式が、私たちの四肢の筋肉と類似することを示しました。これにより、四肢の筋肉をつくる発生様式が、これまで考えられていたよりも古い起源をもつ可能性が示されました。

- Eri Okamoto, Rie kusakabe, Shigehiro Kuraku, Susumu Hyodo, Alexandre Robert-Moreno, Koh Onimaru, James Sharpe, Shigeru Kuratani, Mikiko Tanaka(2017). Migratory appendicular muscles precursor cells in the common ancestor to all vertebrates. Nature Ecology & Evolution 1, 1731-1736.
鰭から四肢への進化に迫るーサメを題材に謎を解き明かすー
鰭から四肢への進化の過程については、原始的な鰭の特徴を持つサメ胚を題材にした研究を展開しています。これまでに、サメ胚の原始的な胸鰭では前側(親指側)が広かったものが、マウス胚の前肢では後側(小指側)が広くなっていること、その原因領域が、前後軸方向のパターン形成を司る Gli3 遺伝子のエンハンサーの機能変化によるものを示しました(Onimaru et al., eLife 2015)。また、数理生物学者の J. Sharpe 氏との共同研究で、サメの鰭の先端の骨パターンは、マウスの指—指間のパターンをつくる Bmp-Sox9-Wnt (BSW)チューリングモデルのパラメーター変化で説明できることを示しました(Onimaru et al., Nature Commun. 2016)。

- Koh Onimaru, Shigehiro Kuraku, Wataru Takagi, Susumu Hyodo, James Sharpe and Mikiko Tanaka(2015). A shift in anterior-posterior positional information underlies the fin-to-limb evolution. eLife 4, e07048.
- Koh Onimaru, Luciano Marcon, Marco Musy, Mikiko Tanaka and James Sharpe(2016). The fin to limb transition as the re-organisation of a Turing pattern. Nature Communications 7, Article number 11582.